

Ijraset Journal For Research in Applied Science and Engineering Technology
Effect of Gibberellic Acid (GA3) on Spore Germination and Reproductive Biology ofChristella dentata (Forssk) Brownsey&Jermy
Authors: Dr. Sandhya Kalera
DOI Link: https://doi.org/10.22214/ijraset.2022.47102
Certificate: View Certificate
Abstract
The climate of Rajasthan is xeric and lacks precipitation but still a good number of pteridophyte, which are shade loving plants and require low temperature and high humidity, grow in different parts of this state. For the present study, the fern Christella dentata was selected and three site were chosen for collection the plant material viz. Nakki lake (Mt. Abu), Sitamata Forest (Baran) and Sitabari (Pratapgarh). In this investigation, the aspects of spore germination and reproductive biology of C. dentata were carried out under the influence of varied concentrations of gibberellic acid GA3. The various populations belonging to the study area exhibited remarkable uniformity in their developmental biology. The mature prothallus is cordate with a prominent midrib bearing rhizoids and sex organs. Studies of effects of GA3 have revealed that 50 ppm of its concentration lead to maximum spore germination and 1- D growth. Maximum 2-D growth was attained at 10 ppm while 20 ppm GA3 could not support gametophytic development to that extent and 100 ppm proved to be lethal. Presence of GA3 in general suppresses the archegonia development and also the development of first leaf of sporophyte.
Introduction
I. INTRODUCTION
In majority of homosporous ferns, the spores develop into hermaphrodite gametophytes. Researches in pteridology tend to inquire into the various aspects of reproductive biology, particularly the gametophytic phase of homosporous pteridophytes. This perhaps is because of the presumption that these gametophytes are commonly or the least potentially hermaphroditic with the initial bearing of antheridia and their continued production thereafter during the production of archegonia. This condition favours intra-gametophytic selfing which leads to homozygous sporophytes (Verma, 2002).
Studies of sex expression of ferns were taken up by Dopp (1950) and he was the first to discover the occurrence of an antheridium inducing substance in fern gametophyte. Pringle, Nag & Braun (1960) and Pringle (1961) named the substance as antheridiogen A. Dopp (1959) and Naf (1958) reported that this substance is effective in inducing antheridium development an all the ferns. Naf (1956) demonstrated that the antheridium formation is controlled by different types of antheridiogens in the various groups of polypodaceous ferns and in particular studied the control of antheridium formation in Anemiaphyllitidis and Lygodium japonicum.
Verma (1989) gave a comparative assessment of the diversity in sex expression and mating system in nine Indian species of Cheilanthes. Yadav (1979) has investigated reproductive biology of some homosporous ferns of Rajasthan. Similarly, Sharma (1979), studied the reproductive biology of some more ferns of Rajasthan which include Adiantum capillus-veneris, A. caudatum, A. lunulatum, Actiniopteris radiata and Cheilanthesfarinosa.
Nutrients as well as other physical and chemical factors, such as light, pH, physical state of medium and plant growth regulators, affect all the processes involved in the growth and development of prothallus (Miller, 1968).
II. MATERIAL & METHODS
For the present investigation spores were collected from the three chosen sites(Site I- Nakki Lake area, Mt. Abu, Site II- Sitabari, Kelwara, Baran, Site III- Sitamata Forest, Pratapgarh). Spores were sprinkled on the culture medium (Knop’s solution) supplemented with 1 ppm of Nitsche’s trace elements and 10 ppm of Ferric citrate.
Spore germination experiments were carried out in culture chamber. The temperature of the culture chamber during experimental studies was kept at 24? to 26?C. Two replicates were set for each treatment in the petri dishes and the relevant data is based on counts of 100 spores per petri dish. A control petri dish was invariably kept along with the set of experiment.
III. OBSERVATION
In the present study effect of the plant growth regulator gibberellic acid GA3 on spore germination and gametophyte development has been studied. Even the chronology of sex expression and mating systems in Christella dentata have been described in this paper. The various populations belonging to the study area exhibited remarkable uniformity in their developmental biology.
A. Spore Germination
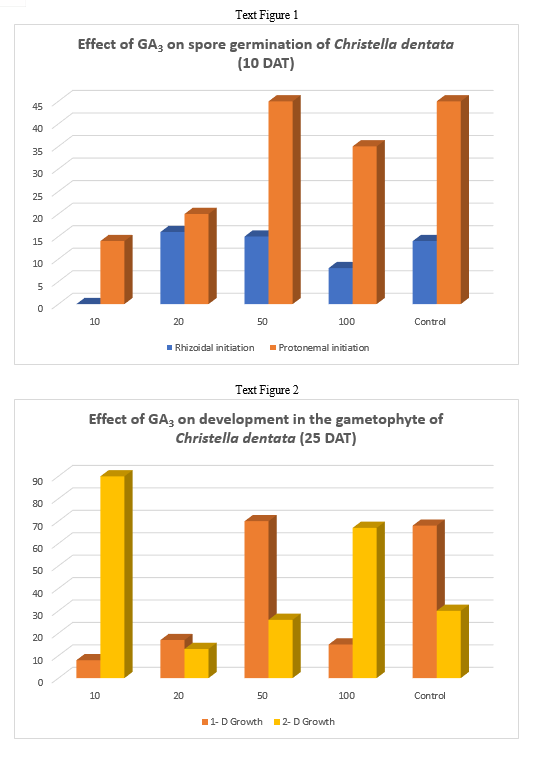
In the present work bursting of exine has been taken to be the first step of spore germination, rhizoid initiation as the next and protonemal initiation as the last step of this process. In the present investigation different concentrations of Gibberellic Acid were added to the culture medium for the study of its effect on the process.
Observations were taken 10 days after treatment. GA3 caused both rhizoidal and protonemal initiation at all concentrations except at 10 ppm. The 10 ppm concentration of GA3 inhibited rhizoidal initiation. Even the control exhibited only but seemingly the percentage of rhizoidal initiation was reduced by 10 ppm of GA3. Maximum rhizoidal initiation was observed at 20 ppm concentration and maximum protonemal initiation was observed at 50 ppm GA3 concentration and in the control.
B. Gametophyte Development
In Christella dentata 1 dimensional growth of gametophytes attaining 2 -dimensional growth was observed in all the concentrations of GA3. Maximum 1 - dimensional growth of gametophytes was recorded at 50 ppm and control. However, in presence of GA3, spathulate and ameristic gametophytes with stout rhizoids were also observed as an abnormal feature.
The maximum 2 - dimensional growth was observed at 10 ppm GA3. Surprisingly 20 ppm GA3 could not support gametophytic development as much as 50 ppm. Full growth of gametophytes was observed nearly 40 days after treatment in all the concentrations except 100 ppm and 20 ppm, which somehow became lethal for further gametophytic growth.
C. Sex Expression
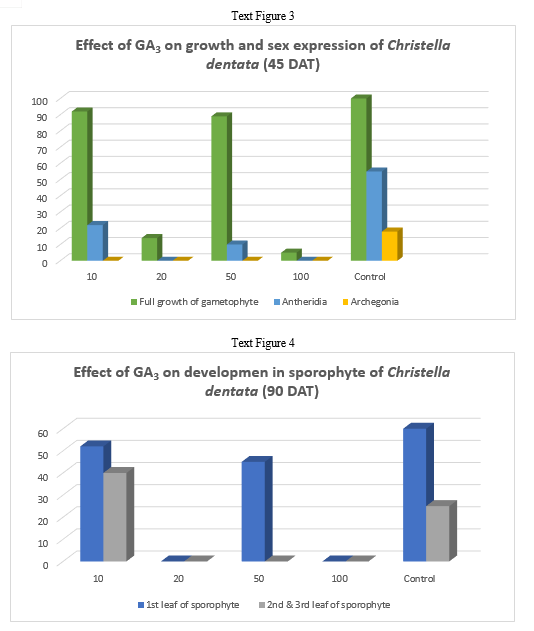
In Christella dentata sex organs were initiated 45 days after treatment. The 10ppm setup was the first to show antheridial development I about 22% cordate gametophytes. The 50 ppm GA3 concentration supported antheridial formation in just 10% gametophytes. However, the antheridial initiation, though delayed a little was highest in the control which even had a few archegonia on the upper half of the cushion.
D. Sporophyte Formation
In Christella dentata, sporophytes developed apogamously nearly 60 days after treatment. Sporophytes were borne on the central region just below the apical notch. Maximum number of prothalli attained hermaphrodite condition in the control. Presence of GA3, perhaps, suppresses the development of archegonia in Christella dentata populations.
Table 1: Effect of GA3 on development and sex expression in gametophyte of Christella dentata (No. of samples studied = 100)
|
S.No. |
Days After Treatment (DAT) |
Parameters |
GA3 Conc. in ppm |
Control |
|||
|
10 |
20 |
50 |
100 |
||||
|
1. |
10 |
Rhizoidal initiation Protonemal initiation |
- 14 |
16 20 |
15 45 |
08 35 |
14 45 |
|
2. |
25 |
1-D growth 2-D growth |
08 90 |
17 13 |
70 26 |
15 67 |
68 30 |
|
3. |
45 |
Full growth of gametophyte Sex expression
|
92
22 - |
14
- - |
89
10 - |
05
- - |
100
55 18 |
|
4. |
60 |
Sex of prothalli
|
59 12 - |
- - - |
49 07 - |
- - - |
65 30 - |
|
5. |
90 |
First leaf of sporophyte 2nd& 3rd leaf of sporophyte |
52 40 |
- - |
45 - |
- - |
60 25 |
Conclusion
Aspects of spore germination, gametophyte development and reproductive biology carried out for present study relates to a study of these processes in response to different concentrations of gibberellic acid (GA3) on spores of Christella dentata. The pattern of spore germination and gametophyte development in C. dentatawas that of Vittaria type and Drynaria type respectively, as described by Nayar& Kaur (1971). Under the influence of GA3 the percentage of rhizoidal initiation decreased considerably at all concentrations. Optimum concentration for gametophytic growth was recorded to be 10 ppm. The gametophytes of homosporous pteridophytes are commonly, or least potentially, hermaphroditic with the initial bearing of antheridia and their continued production thereafter during the formation of archegonia. Such a condition has been taken to be highly permissive in intra-gametophytic selfing that lead to completely homozygous sporophytes.
References
[1] Dopp, W. 1950. Eine die AntheridienbildungbeiFarnenforderndeSubstanz in den Prothallien von Ptreidium aquilinum (L.) Kuhn. Ber. Deut. Bot. Ges. 63: 139-147. [2] Dopp, W. 1959. Uber einehemmende und einefordendeSubstanzbei der Antheridienbildung in den Prothallein von Ptreidium aquilinum Ber. Deut. Bot. Ges. 72: 11-24. [3] Miller, J.H. 1968. ‘Fern gametophytes as experimental material.’ Bot. Rev. 34: 361-440. [4] Naf, U. 1956. The demonstration of a factor concerned with the initiation of antheridia in Polypodiaceous ferns. Growth. 20:91-105. [5] Naf, U. 1958. On the physiology of antheridium formation in the bracken fern Ptreidium aquilinum (L.) Kuhn. Physiol Plantarum. 11: 728-746. [6] Nayar, B.K. & Kaur, S. 1970. Gametophytes of homosporous ferns. Bot. Rev. 37: 295-296. [7] Pringle, R.B. 1961. Chemical nature of antheridiogen-A, a specific inducer of the male sex organ in certain fern species. Science. 43:133-284. [8] Pringle, R.S., Nag, U. & Braun, A.C. 1960. Purification of a specific inducer of the male sex organ in certain fern species. Nature. 186: 1066-1067. [9] Sharma, P. 1979. Cytotaxonomic and experimental studies on some leptosporangiate ferns of Rajasthan. Ph.D. Thesis, University of Rajasthan, Jaipur. [10] Verma, S.C. 1989. Overt and Covert mechanism of intergametophytic mating in homosporous ferns. In: Plant Science Research in India. (Eds.). Trivedi, M.L., Gill, B.S. & Saini, S.S. New Delhi: Today and Tomorrow’s Printers and Publishers, pp. 285-300. [11] Verma, S.C. 2002. “Gametophytic Reproductive Biology of Homosporous Ferns: An Assessment.” In: Advances in Pteridology. (Ed.) Trivedi, Pravin Chandra. Jaipur (Rajasthan): Pointer Publishers, pp. 48-80. [12] Yadav, A.K. 1979. Phytogeography, distribution, cytology and developmental studies of some Rajasthan ferns. Ph.D. Thesis, University of Rajasthan, Jaipur.
Copyright
Copyright © 2022 Dr. Sandhya Kalera. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

Download Paper
Paper Id : IJRASET47102
Publish Date : 2022-10-17
ISSN : 2321-9653
Publisher Name : IJRASET
DOI Link : Click Here
 Submit Paper Online
Submit Paper Online