

Ijraset Journal For Research in Applied Science and Engineering Technology
Effect of Salinity on Some Physiological Parameters in Tomato
Authors: Kada Mokhtar Derkaoui, Prof. Mohamed Sahnoune, Moulay Belkhodja
DOI Link: https://doi.org/10.22214/ijraset.2022.40445
Certificate: View Certificate
Abstract
Tomatoes are an important plant in terms of human consumption in many countries of the world. The salinity of soils and irrigation water is one of the limiting factors of plant productivity and agricultural yield. The present work consisted of evaluating the physiological and enzymatic behaviour of the four tomato varieties, Marmande, Aicha, Henz1573 and Cerise, subjected to increasing concentrations. The results obtained showed that the sensitivity of the four varieties was observed in a variable way from one genotype to another. Indeed the levels of chlorophyll a, b and total have been reduced under the effect of saline stress, while the concentration of proline has increased. The content of 1-aminocyclopropane-1-carboxylic acid (ACC) determined in the leaves increased significantly as a function of salinity levels and genotypes. The rate of ethylene production in tomato petioles was a function of salinity levels, leaf age and genotype.
Introduction
I. INTRODUCTION
Physiological dryness, a term often attributed to osmotic stress in salinity conditions, subjects the plant's tissues to a certain dehydration. Salinity can be natural or induced by agricultural activities such as irrigation, where more than 30% of water intended for irrigation is loaded with salt, resulting in a reduction in the growth and yields of sensitive varieties (Bouzid, 2012) [1] or the use of certain types of fertilizers.
In the world Each year, the areas lost due to soil salinity vary around 20 million hectares including 10 million arable land, or 23% of cultivated land, one billion ha is threatened, including 3.2 million ha in Algeria (Kaci and al, 2012) [2]. The regions of the Mediterranean Basin are characterized by very restrictive pedoclimatic conditions: aridity, salinity, extreme pH, soils deficient in organic matter and minerals, summer drought and strong carbonation of most soils. In Algeria, 80% of cultivated land is located in climates ranging from semiarid to arid and, each year, thousands of hectares are removed from agropastoral use due to desertification combined with salinity (Domergue, 2006) [3]. Ion toxicity resulting from the accumulation of specific ions such as Na+ and Cl– in the cytoplasm or apoplast interferes with the metabolic functions of the plant (Levitt 1980) [4]. Understanding the metabolic processes responsible for salinity tolerance and the factors that control their expression is fundamental to any approach aimed at improving the performance of tomato cultivation under salinity conditions. Saline stress influences growth through many facets of metabolism, such as nutrient absorption (Grattan and Grieve 1993) [5] and their distribution within the plant (Grattan and Grieve 1999) [6], impaired photosynthesis (Belkhodja 1993) [7] and respiration, protein and nucleic acid synthesis (Ben-Hayyim and al. 1989) [8], accumulation of organic solutes, enzyme activity (Dubey and Rani 1990) [9], hormonal balance and water availability (Hamza 1980) [10].
This article discusses some physiological and biochemical characteristics of salinity sensitivity by examining the responses of different tomato genotypes to increasing levels of salinity. The list of metabolites influenced by salinity, as reported in the literature, is long. In our case, we will limit ourselves to metabolites that have shown significant variations in other species. Thus, we will examine the effect of salt on chlorophyll content, proline accumulation, Na+/K+ selectivity ratio, ethylene production and 1-aminocyclopropane-1- carboxylic acid (ACC) concentration. Indeed, genotypic variations under saline stress have been reported for the Na+/K+ ratio (Gorham 1990; Mekkaoui and al. 1994) [11] [12], for proline accumulation (Rhodes and Handa 1989; Mekkaoui and al. 1994) [13] [12], for chlorophyll (Tewari and Singh 1991; Idihia 1995) [14] [15] and for ethylene (Khan et al. 1987; Prusinski and Khan 1990) [16] [17].
Genotypic variations under water stress have been reported for ACC (Benichou and al. 1993) [18]. Thus, the specific objective of this study is to look for one or more criteria to highlight the sensitivity to salt of tomatoes. To do this, different genotypes of different levels of salinity tolerance were examined in controlled experiments.
II. MATERIALS AND METHODS
A. Preparation of Plant
The sowing of tomato seeds was carried out in the nursery and then the transplanting of the seedlings was carried out in a pot in a greenhouse Experimental protocol Location of the test The trial was conducted at the Faculty of Natural and Life Sciences in Tiaret; under semi controlled conditions (plastic greenhouse). The plant material used The varieties of tomatoes tested in this test are:
Marmande, Heinz 1573 Cerise and Aicha. Experimental device The test shall be conducted in plastic cylinders (P.V.C) with a diameter of 10 cm and a height of 100 cm, each filled with a substrate composed of a mixture of well-decomposed organic matter, sand and earth according to the respective proportions of 1.8.1. This device has three blocks; each block consists of 12 cylinders with four varieties of tomato repeated with three repetitions. three saline concentrations are applied with 75, 145 and 200 mM of NaCl respectively. For the control lot is irrigated with tap water (not salted). The experiment was conducted according to a random experimental device with four treatments. Sowing is carried out in the nursery on March, 2013 at the rate of two seeds per pot at a depth of 1 cm.
Transplanting is carried out one month after sowing at the rate of one plant per cylinder. The seedlings are watered with a nutrient solution until the six-leaf stage when saline treatment has begun. Phytosanitary treatment Before the start of the experiment, the substrate used for the experiment was treated with an insecticide (karate) and a fungicide (Anvil5Sc). Irrigation Every 48 hours’ irrigation is maintained at capacity in the field by the contribution of 250 ml of water for all lots; to calculate the watering dose, the weight of the pot is subtracted after twenty-four hours of wiping from the weight of the pot saturated with water. From transplanting until the application of saline stress, irrigation is provided with a nutrient solution, diluted in distilled water at a rate of 2g/l.This solution contains balanced nutrients to strengthen the vigor of plants, stimulate rooting, improve the quality of crops and help the plant recover quickly in the event of a climatic accident. Methods and measurements carried out The measurements were carried out one month after the application of the saline solution and concerned the morphological parameters; physiological of the aerial part.
*FM: Fresh matter
B. Measurement parameters
- Chlorophyll Content: The method used is that of Lichtenthaler and Welburm (1983) modified by Porra (2002) [19], it consists in grinding a portion 100 mg of sheet in a porcelain mortar in 8ml of acetone diluted to 80%. The grind is filtered into a test tube using Watman Paper No. 22. Then the volume of the tube is completed to 10 ml by adding diluted acetone. The absorbances were read by the spectrophotometer at wavelengths 645 nm and 663 nm. The absorbances were converted according to the authors in order to quantify the content of chlorophyll a, chlorophyll b and total chlorophyll in 1.
Ug/g MF, using the following formulas:
Chla=(12.21x663nm)-(2.81x645nm) .
Chl b = (20.13x645 nm)-(5.03x663 nm) .
Chl t= Chl a+ Chl b .
2. Proline Content: Proline was determined using the method of Idihia (1995) [15]. Samples of leaves and petioles of the same leaf stage (4th leaf from the base) and fruits of the 1st bunch were taken from 4 plants at random per elementary plot 45 days after the start of saline treatments.
3. Determination of Mineral Matter Composition : The plant was harvested after the treatments as defined and separated from the roots, and dried in a forced air oven at 65?C for 4 days for the determination of the dry weight (DS). The dried material was digested with HNO3:HClO4 (2:1, v:v). Concentrations of K+, Mg2+ and Na+, Cl- were determined by ICP-MS using an Iris Intrepid ICP spectrometer (Thermo Electron Corporation, Franklin, USA).
4. Acc(1-Aminocyclopropane-1- carboxylic acid) Content : ACC levels were determined by the indirect assay method of converting ACC to ethylene (Lizada and Yang 1979) [20]. The ethylene was determined by gas chromatography with flame ionization detection. The separation was made by the Poropak Q column (1.5 m in length) mounted in the type chromatograph (Delsi 330). These assays were carried out on samples of leaflets, petioles (4th leaf from the apèxe).
5. Ethylen content: Ethylene accumulation was measured as described by Shibli and al. (1997a) [21] using gas chromatography. Prior to sampling, flasks were capped for 3 h with rubber caps and a 1 ml sample of headspace gasses was withdrawn from each flask through the septum with a hypodermic syringe, and injected into a GC (Perkin Elmer, Norwalk, CT, USA) equipped with flame ionization detector and an activated alumina column. Oven temperature was 80C and detector and injector temperature was 200C. Helium was used as a carrier gas.
6. Aerial Dry Matter : Dry above-ground biomass was determined using a precision balance. The fresh weight was directly determined after the recovery of the organ. The dry peas were obtained after the passage of the organs 48 hours in the oven at 80 ° C.
7. Statistical analysi : Statistical processing was performed by Excel, Version 2016 and the parameters recorded were subjected to two-way analysis of variance. Comparison means were performed by ANOVA test at the significant level of 0.05.
III. RESULTS AND DISCUSSIONS
A. Effet de la salinité sur la chlorophylle
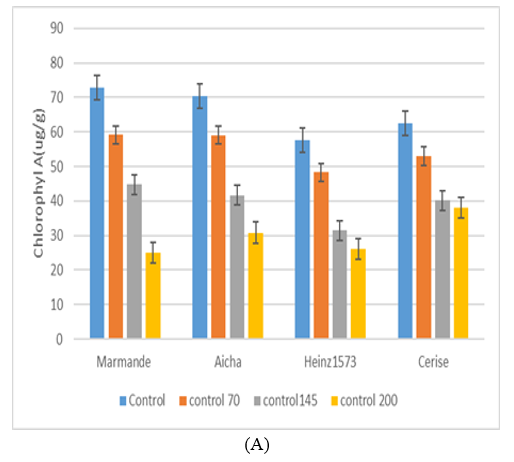
- Chlorophyll A : Tomato varieties Marmande, Aicha, Henz1573 and Cerise showed a significant decrease from their control and showed percentage reductions of 59.18, 59.09, 48.25 and 52.96 ug/g respectively (Fig. 1). When the stress is severe (200mM), the chlorophyll content A is even more affected, especially in the case of Marmande and Aicha whose percentage reduction is 65% and 56% respectively compared to that of the control. The statistical study shows that there are significant differences between the varieties studied (p<0.05) and shows that under severe stress, the Cerise genotype has a better salt tolerance with a percentage of 38% compared to the controls, followed by Henz1573 with 54% reduction, and keep a chlorophyll content A quite high.
- Chlorophyll B : In this case (Fig.1), tomato plants Marmande, Aicha, Henz1573 and Cerise recorded a percentage reduction of 6, 35, 25, and 33% respectively compared to the control. At the level of the saline concentration 150 mM, the Aicha variety recorded a greater reduction in chlorophyll B with 63% the other genotypes Hen1573, Cerise and Marmande showed reductions of 51I% and 45% respectively. Thus the other cultivars show considerable reductions in this content compared to the controls for Marmande, Aicha and Henz1573 with percentages of 74%, 75% and 72% respectively. In addition, our results show that chlorophyll (B) content is more sensitive to the effect of saline stress than chlorophyll (a).
- Chorophyll a+b : Fig.1 for this parameter shows that Aicha and Marmande are the varieties richest in total chlorophyll, in the absence of saline stress. When these plants are subjected to moderate stress 70mM, the percentage reduction compared to the control varies from 14 to ,23,20 and 21 in the Plants Marmande, Aicha, Henz1573 and Cerise. In case of severe stress at 200mM, Cerise and Aicha keep a fairly high chlorophyll content compared to the others with 41.62 ug/g MF and 44.89ug/gMF respectively with a reduction rate of 52% and 63%. On the other hand, Marmande and Henz1573 have a percentage reduction compared to the control of 68% and 61%. These results differ from those of Sivtsev (1973) [28] on tomato, Tewari and Singh (1991) [14] on lentil (Lens esculenta M); but are substantially in line with the results of Sharaf and al. (1990) [29] on tomatoes. The latter report that moderate salinity increases the amount of chlorophyll a and total chlorophyll. Sodium chloride has less of an impact on the chlorophyll b biosynthesis pathway (Tewari and Singh, 1991) [14].
B. Effect of Salinity on Proline Accumulation
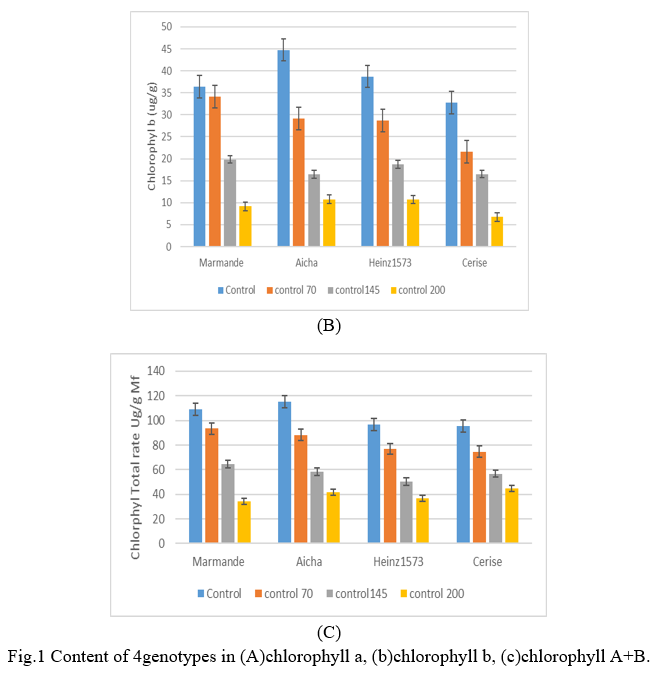
ANOVA showed the significant effect of salinity on proline production regardless of genotype and organ and that proline accumulation increased in the leaves of all four tomato genotypes under the effect of saline stress. The average results obtained in Fig.2, show the content of the proline produced in the leaves of the four tomato lines subjected to the different saline concentrations. There is an increase in the proline content in stressed plants compared to the control, the highest levels were recorded in plants under severe stress 2000mM in all lines. Where the Marmande line, displayed the highest content with 0.75mg/ml representing an increase of 243% compared to the control. In terms of absolute value, the Cerise variety accumulated the minimum of proline under saline stress with 0.47mg/ml, which represents, a rate of increase of 215% while the tomato Aicha and henz1573 showed an intermediate accumulation with respectively the values 0.49 mg/ml (a lower rate of increase with 186%) and 0.51 mg/ml (its rate of increase is 198%). At the level of the stressed batch at 145mM, the percentage of proline accumulation was higher in Marmande (162%) with an amount of 0.57mg/ml and for the lowest amount is recorded in Cerise with 0.34mg/ml (133%). Depending on the genotype, the accumulation of intermediate proline is displayed by Aicha and henz1573 with the following proline amounts in order of 0.43mg/ml (151%) and 0.44mg/ml (155%). We notice in the saline concentration 70mM, the Marmande variety displays a very important amount of proline produced which is of the order of 0.45 mg / ml and which is 108% compared to the control. On the other hand, the lowest amount is recorded by the Cultivar Aicha which is 0.36mg / ml with a rate of increase compared to the control of 23%. The intermediate amounts are presented by the Henz1573 variety with 0.33mg/ml (92%) and the Cherry variety with 0.27mg/ml (83%). Depending on the organ and genotype, the accumulation of proline, which in our case reached values 2 times higher than the control value, is similar to the concentrations measured by Huer (1993) [22] in tomatoes, but very far from the values cited by Rhodes and Handa (1989) [13].
The latter report that tobacco cells adapted to 428 mM of NaCl accumulate 80 times more proline than unsuitable cells. The accumulation of proline is attributed to the stress inhibitory effect on its oxidation in mitochondria (Sell and Koepp 1981) [23], as well as on its incorporation into proteins (Hsiao 1973) [24].
The neosynthesis of the proline would be triggered by the loss of turgor due to salinity. This activates a series of complex events correlated with the level of stress, the tolerance of the plant and its growth (Huer 1993) [22].
While several authors have linked proline accumulation to plant tolerance to salinity, (Rhodes and Handa 1989; Delauney and Verma 1993) [13], [25], others report the absence of this correlation (PerezAlfocea and al. 1993; Guerrier 1998) [26], [27]
C. Evolution of the selectivity ratio K+/Na+
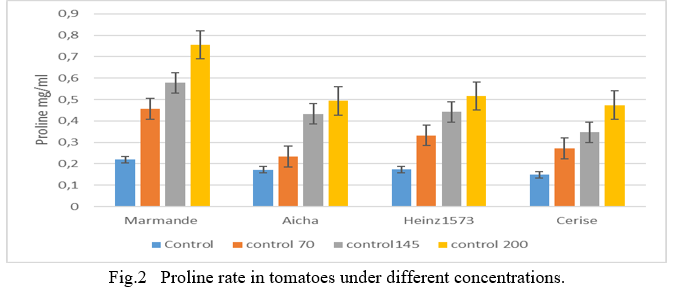
The average results obtained in the fig show the ratio Na+/K+ decreased in the leaves of the four lines subjected to the different saline concentrations. There is decrease in stressed plants compared to the control, the highest levels were recorded in plants under severe stress in all lines, the highest content is recorded at the concentration 200 mM with 1,54 in the Henz1573 variety (Fig.3.). The lowest value is displayed by The Marmande genotype with 0,93. The other cultivars have intermediate values, in particular Aicha with 1,14 and Cerise with 0,99. At the level of the treatment 145mM records a high value in Henz1573 is 0,75 which constitutes an increase of 76% with the control and the lowest value is 0,52 in Cerise with a raise rate of 82% of the control. It can be seen that Henz1573 at the 70 mM treatment level with a value of 0,93its the biggest one, the lowest value is noticed in Marmande with a value of 0,69. The other varieties Aicha and Cerise recorded intermediate leaf- ratio of the order of 0,87 and 0,79 respectively. So the selectivity ratio K+/Na+ decreased in the leaves of all genotypes following the increase in salinity (Fig.3); which shows that the selectivity in the leaves is in favor of potassium. Cramer and al. (1985) [30] demonstrated that in the presence of high concentrations of NaCl, Na+ displaces Ca++ from plasmaleme of root cells which leads to increased membrane permeability and causes K+ efflux and impaired K+/Na+ selectivity ratio. This suggests that plants that successfully grow in saline environments are those that maintain a higher K+/Na+ ratio in their cytoplasm than in the rhizosphere. With high concentrations of Na+ in the leaf apoplast and/or vacuole, plant cells have difficulty maintaining low cytosolic Na+ and, perhaps as importantly, low Na+/ K+ ratios cited by Gorham (1990) [11]. In addition, Cuartero and Fernandez-Munoz (1999) [31] linked the tolerance of tomato plants to salinity to their ability to control Na+ in adult leaves while maintaining a low concentration of this element in young leaves.
D. Effect of Salinity on 1-aminocyclopropane-1-carboxylic acid (ACC)
Saline stress caused the concentration of 1-aminocyclopropane-1-carboxylic acid (ACC) to increase in tomatoes. This determined concentration in the leaves of tomato plants exposed to all three levels of salinity increased significantly with the increase in salinity levels (Fig.4). The content of this organ in ACC at the most stressed level 200mM was higher than at the non-stressed level. Thus, the ACC values in the leaves increased from 7.74 nmol g–1 mf (minimum value under non-stress conditions) to 17.62nmol g–1 mf (maximum value under stress conditions); in the henz1573 variety and from 6.87 to 16.64nmol g–1 mf at the cherry level. The lowest ACC values were recorded in Aicha and Marmande with 10.32 nmol g–1 mf and 11.60 nmol g–1 mf, respectively. Fuhrer (1982) [32] reported that stimulation of the induction of ACC synthase by stress is responsible for the accumulation of ACC. Indeed, the treatment of tomato plants exposed to saline stress with ACC synthase inhibitors such as AOA (alpha-aminooxyacetic acid) suppressed this accumulation (Jones and El-Abd 1989) [33].
Hoffman and Yang (1980) [34] explained the lesser accumulation of ACC in susceptible varieties by their inability to convert s-adenosylmethionine (SAM) to ACC. In the batch at 145mM, the concentration of ACC was very low, detected in the Cherry variety followed by Marmande with 9.14 nmol g–1 mf and 10.62 nmol g–1 mf respectively. The Henz1573 verieté had the greatest value with 16.32 nmol g–1 mf and also the Cherry variety with 15.96 nmol g–1 mf. At the concentration level at 70mM, the lowest amount of ACC is recorded by the cultivar Henz1573 followed by Marmande with values respectively 6.78nmol g–1 mf and 9.10 nmol g–1 mf. the most important value is displayed by the variety Henz1573 13.96 nmol g–1 mf and also the variety Cherry with 12.98 nmol g–1 mf . Hoffman and Yang (1980) [34] also found very low values and attribute these to the inability of these tissues to convert SAM to ACC. Furthermore, Larrigaudière et al. (1990) [35] report that the stimulation of ethylene production by stress is probably the result of pre-existing mRNA activation because this synthesis can be avoided by cycloheximide (translation inhibitor) but not by cordycepine (transcription inhibitor).
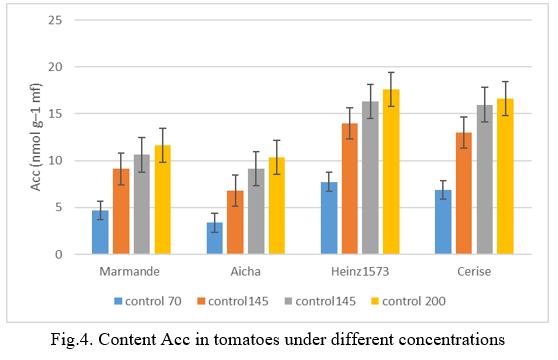
E. Effect of salinity on ethylene production
The production of ethylene in tomato leaves increased significantly with the increase in the level of salinity in the root environment of the plant (Fig5). The values for ethylene are slightly higher than the values reported by Abou Hadid and El-Beltagy (1986) [36] in tomatoes, but much lower than those found in wheat by Narayane and al. (1991) [37]. The latter believe that monocots and broadleaf weeds can have a fundamental difference from ethylene especially if ethylene is related to processes such as abscission which is specific to dicots. This ethylene production also differed significantly between genotypes. The henz1573 variety (Fig.5) recorded the highest amount of ethylene, both in salinity and in the absence of salinity. The Aicha variety had the lowest ethylene value under non-stress conditions and remained below the rates of other genotypes even with the imposition of saline stress. The level of ethylene and the dry matter accumulated by the different genotypes (El-Iklil and al. 2000) [38].
This suggests links between the rate of ethylene production and salinity tolerance in tomatoes. The results also show that the rate of ethylene production depends on the age of the leaf where the petioles are taken. It was, according to the salinity level 200mM in henz1573 of 11.93 nmol g–1 mf on the second leaf and 13.66 nmol g–1 mf on the fourth leaf. Aicha presents on the second sheet the amount of ethylene which is 5.8625 nmol g–1 mf and on the fourth sheet a value of 5.01 nmol g–1 mf. The other varieties record intermediate quantities. (Fig.3). Similar results were found by Abu Hadid and El-Beltagy (1986) [36] who reported that ethylene is more important in adult leaves than in young leaves under stress.
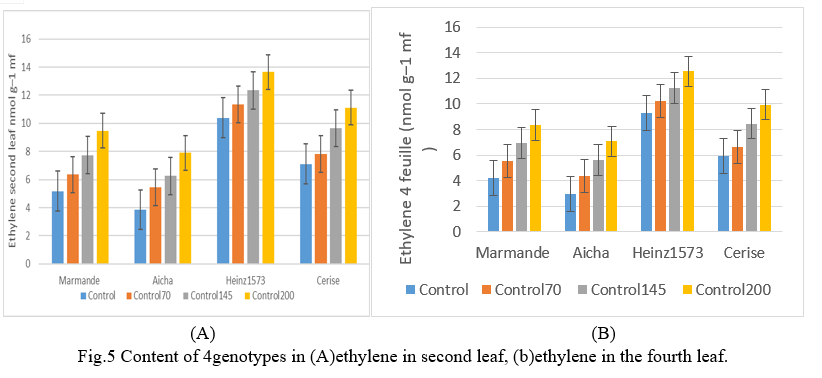
F. Salinity Effect on Aerial dry Matter
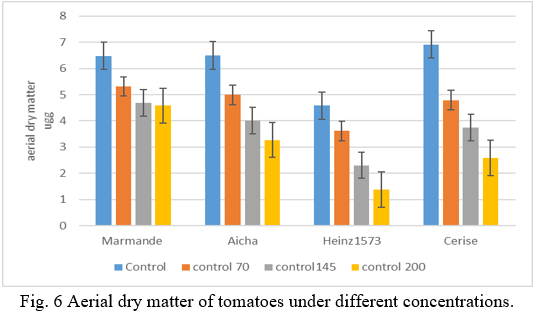
The results in (Fig.6) show that dry above-ground biomass is affected in all lines studied. A decrease in all lines for control conditions. (Fig.6) shows that the highest dry above-ground biomass is recorded in stressed plant levels with 70mM, specifically the Marmande and Aicha lines with 5.31g and 4.99g respectively, compared to controls.
The lowest values are recorded by Henz and cherry with 3.61g and 4.79g respectively. For plants subjected to a stress of 150mM of which values have been recorded that vary between 4.69g at the level of La Marmande stressed and 4.01 at the level of the line Aicha stressed. Note that the small value is displayed by Henz1573gavec 2.30g followed by Cerise with 3.74g. The application of a more severe stress of 200mM causes a high decrease in dry matter at the line level, especially the Henz1753 variety records the lowest amount with 1.37g followed by the Cherry variety with 2.58 g. While the Marmande variety records the great value with 4.58 g followed by Aicha with a value of 3.27g. The results of the analysis of variance revealed a significant difference between tomato lines subjected to different saline concentrations and this parameter (P<0.05). According to Ben Ahmed and al. (2008) [39], the depressive action of salt is manifested by a reduction in the production of dry matter of the various organs of the plant. It is also manifested by reduced plant height (Singh and Prasad, 2009) [40].
IV. ACKNOWLEDGMENT
The authors express their gratitude to Pr. Ahmed Adda, director of the vegetal biology Laboratory, biology and agriculture institute ‘s staff of Tiaret for the reception and hospitality. They also want to thank the beekeepers for their collaboration.
Conclusion
IV. CONCLUSIONS The efficiency of a criterion to express sensitivity or tolerance depended on several factors: 1) The level of salinity: the selectivity ratio Na+/K+ was more discriminating at moderate salinity than at high salinity. 2) The organ analyzed: the effect of salinity on the accumulation of proline as a safety criterion was more evident in the leaves. 3) The age of the organ analyzed: the production of ethylene was greater in the young leaves than in adults. 1-Aminocyclopropane 1-carboxylic acid (ACC) is the direct precursor to ethylene, a plant hormone. ACC is synthesized from S-adenosyl-L-methionine (SAM) by ACC synthases (ACS) and then oxidized to ethylene by ACC oxidases (ACO). Ethylene is a simple molecule with two carbon atoms with profound effects on plants. There are numerous synthesis studies covering all aspects of the biology of ethylene in plants, including its biosynthesis, signaling and physiology. This is only a logical consequence of the fascinating nature of this plant hormone. Its biochemical precursor, 1-aminocyclopropane-1-carboxylic acid (ACC) is also a fairly simple molecule, but perhaps its role in plant biology is seriously underestimated. Finally, this study highlighted the existence of some genotypic variability for the metabolites analyzed. The latter may therefore constitute potential criteria for characterizing salinity tolerance in tomatoes. However, a thorough study of the heritability of these traits remains essential in any approach to the choice of salinity tolerance criteria.
References
[1] Bouzid S, Rahmoune C., 2012. Enhancement of Saline Water for Irrigation of Phaseolus vulgaris L. Species in Presence of Molybdenum.Procedia Engineering, 33, 168 –173p. [2] Kaci S., Bissati S. et Djerroudi O., 2012. Effet d’un stress salin sur la réponse minérale d’Atriplexcanescens (PURSH) NUTT., Revue des Bioressources. Université KASDI MERBAH Ouargla. Algérie 48-58p. [3] Domergue O., 2006. Diversité des rhizobia associés à Ononis repens: une légumineuse adaptée aux milieux méditerranéens. Mem.ing. Ecole pratique des hautes études. Montpellier, 8p. Abou Hadid, A. F. et El-Beltagy, A. S. 1986. Ethylene production by leaves of various plant species in response to stress imposition. Acta Hortic. 190: 415–422. [4] Levitt, J. 1980. Responses of plants to environmental stress. Vol.II. Water, salt and other stresses. Academic Press, New York, NY. [5] Grattan, S. R. et Grieve, C. M. 1993. Mineral nutrient acquisition and response by plant grown in saline environment. Pages 203–226 dans M. Pessarakli, ed. Handbook of plant and crop stess. MarcelDekker, Inc., New York, NY. [6] Grattan, S. R. et Grieve, C. M. 1999. Salinity-mineral relations in horticulural crops. Sci. Hortic. 78: 127–157. [7] Belkhodja, R. 1993. La fluorescence de la chlorophylle sur l’orge(Hordeum vulgare L.): une possible voie pour le criblage de variétés tolérantes à la salinité et à la sécheresse. Master of ScienceThesis. Instituto Agronomico Mediterraneo de Zaragoza, Spain. [8] Ben-Hayyim, G., Vaadia, Y. et William, B. G. 1989. Proteins associated with salt adaptation in citrus and tomato cells: Involvement of 26 KD polypeptides. Physiol. Plant. 77: 332–340. [9] Dubey, R. S. et Rani, M. 1990. Influence of NaCl salinity on the behavior of protease, aminopeptidase, and carboxy-peptidase in rice seedlings in relation to salt tolerance. Aust. J. Plant Physiol. 17: 215–221. [10] Hamza, M. 1980. Réponses des plantes à la salinité. Physiol. Vég. 13: 69–81. [11] Gorham, J. 1990. Salt tolerance in the Triticeae: K/Na discrimination in Agilops species. J. Exp. Bot. 41: 615–621. [12] Mekkaoui, M., Agbani, M. et Monneveux, P. 1994. Rôle de la sélectivité et de l’accumulation de la proline dans l’adaptation à la salinité de l’orge (Hordeum vulgare L.) et du blé (Triticum durum Desf.). Actes Inst. Agron. Vet. (Maroc) 14: 27–36. [13] Rhodes, D. et Handa, S. 1989. Amino acid metabolism in relation to osmotic adjustment in plant cells. Pages 41–62 dans J. H. Cherry, ed. Environmental stress in plants, biochemical and physiological mechanisms. Springer-Verlag, New York, NY. [14] Tewari, T. N. et Singh, B. B. 1991. Stress studies in lentil (Lens esculenta M.) II. Sodicity induced changes in chlorophyll, nitrate and nitrate reductase, nucleic acid, proline, yield and yield components in lentil. Plant Soil 136: 225–230. [15] idihia F 1995 ,Caracterisation agro-morphologique de la tolerance à la salinité chez l\'orge (hordeumvulgare).Thése de troisieme cycle.Université Cadii Ayyad,Faculté des sciencesSemlalia,Marrakech. [16] Khan A. A., Akbar M. et Seshu, D. V. 1987. Ethylene as an indicator of salt tolerance in rice. Crop Sci. 27: 1242–1247. [17] Prusinski, J. et Khan, A. A. 1990. Relationship of ethylene production to stress alleviation in seeds of lettuce cultivars. J. Am. Soc. Hortic. Sci. 115: 294–298. [18] Benichou M., El Mourid M. et El Boustani, E. 1993. Amélioration de la méthode de l’analyse chimique de l’acide-1-Malonylaminocyclopropane-1-carboxylique (MACC) dans un extrait brut de feuilles de blé. Al Awamia 81: 125–139. [19] Porra, R.J., 2002. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynthesis Research 73, 149–156 [20] Lizada, M. C. C. et Yang, S. F. 1979. A simple and sensitive essay for 1-aminocylopropane carboxylic acid. Anal. Biochem.100: 140–145. [21] Shibli RA, Sawwan J, Swaidat I, Tahat M (2001) Increased phosphorus mitigates the adverse effects of salinity in tissue culture. Communic Soil Sci Plant Anal 32:429– 440. [22] Huer, B. 1993. Osmoregulatory role of proline in water and salt stressed plants. Pages 363–381 dans M. Pessarakli, ed. Handbook of plant and crop stress. Marcel Dekker, Inc., New York, NY. [23] Sell, G. D. et Koepp, D. E. 1981. Oxidation of proline by mitochondri isolated from water stressed maize shoots. Plant Physiol 68: 1058–1063. [24] Hsiao, T. C. 1973. Plant responses to water stress. Ann. Rev. Plant Physiol. 24: 519–570. [25] Delauney, A. J. et Verma, D. P. S. 1993. Proline biosynthesis and osmoregulaton in plants. Plant J. 4: 215–223. [26] Perez-Alfocea, F., Estan, M. T., Caro, M. et Guerrier, G. 1993. Osmotic adjustment in Lycopersicon esculentum and Lycopersicon pennillii under NaCl and polyethylene glycol 6000 isosmotic stress. Plant Physiol 87: 493–498. [27] Guerrier, G. 1998. Proline accumulation in salt-treated tomato: different proline precursors in Lycopersicon esculentum and Lycopersicon pennillii. J. Plant Nutr. 21: 505–513. [28] Sivtsev, M. 1973. Photochemical activity of chloroplast and bound strength complex in cultured plants during action of salinization and biologically active compounds Fizol. Rast. 20: 1176–1181. [29] Sharaf, A., Labib, S. et El Massry, R., 1990. Effect of kinetin on the biochemical constituents of tomato plants under different levels of salilnity. Zagazig Journal of Agricultural Research (Egypt).12: 417-441. [30] Cramer, G. R., Lauchli, A. et Polito, V. S. 1985. Displacement of Ca2+ by Na+ from the plasmalema of root cells. A primary response to salt stress. Plant Physiol 79: 207–211. [31] Cuartero, J. et Fernandez-Munoz, R. 1999. Tomato and salinity. Sci. Hortic. 78: 83–15. [32] Fuhrer, J. 1982. Ethylene biosynthesis and cadmium toxicity in leaf tissue of beans (Phaseolus vulgaris L.). Plant Physiol. 70: 162–167. [33] Jones, R. A. et El-Abd, S. O. 1989. Prevention of salt induced epinasty by alpha-aminooxyacetic acid and cobalt. Plant Growth Reg. 8: 315–323. [34] Hoffman, N. E. et Yang, S. F. 1980. Changes of 1-aminocyclopropane-1-carboxylic acid content in repining fruits in relation to their ethylene production rates. J. Am. Soc. Hortic. Sci. 105: 492–495. [35] Larrigaudière, C., Latché, A., Pech, J. C. et Triantaphylidès C. 1990. Short-term effect of ?-irradiation on 1-amonocyclopropane1-carboxylic acid metabolism in early climatric cherry tomatoes.Comparison with wounding. Plant Physiol. 92: 577–581. [36] Abou Hadid, A. F. et El-Beltagy, A. S. 1986. Ethylene productionby leaves of various plant species in response to stress imposition. Acta Hortic. 190: 415–422. [37] Narayana.I,S.Lalonde,and H.S.Saini.1991.Water-stress-induces ethylele production inwheat plant physiol.96:406-410. [38] El-Iklil, Y., Karrou, M. et Benichou, M. 2000. Salt stress effect on epinasty in relation to ethylene production and water relations in tomato. Agronomie 20: 399–406. [39] Ben Ahmed H, Arafet M, Zid E. 2008. Tolérance à la salinité d’une Poaceae à cycle court : la sétaire (Setaria verticillata L.). C.R. Biologies, 331:164- 170. [40] Singh A, Prasad R. 2009. Salt stress growth and cell bound enzumes in Archis hypogea L. seedling. I.J.I.B., 7(2): 107- 123.
Copyright
Copyright © 2022 Kada Mokhtar Derkaoui, Prof. Mohamed Sahnoune, Moulay Belkhodja. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

Download Paper
Paper Id : IJRASET40445
Publish Date : 2022-02-21
ISSN : 2321-9653
Publisher Name : IJRASET
DOI Link : Click Here
 Submit Paper Online
Submit Paper Online