

Ijraset Journal For Research in Applied Science and Engineering Technology
Thermal Soaring and Control Surface Dynamics of an Eagle and the Hummingbird’s Flapping Flight Aerodynamics
Authors: Prantik Dutta, Samruddhi Chaugule
DOI Link: https://doi.org/10.22214/ijraset.2022.47148
Certificate: View Certificate
Abstract
This review tends to shed light on the eagle’s aerodynamic attributes along with its inbuilt structural control surfaces that facilitate its high maneuverability. Apart from this, few aspects of the eagle’s wing morphing techniques, its aerofoil, and its aerodynamic stability are also being highlighted. The aim is to highlight the bio-mimicable traits of an eagle which can be incorporated into bionic UAVs. This work acts as a base for current and future works involving slotted wingtips and bionic control surfaces. The traits discussed are being used to design mechanical control surfaces and wingtips that resemble the eagle’s slotted wings and control surfaces. These works when combined with compliant mechanisms can help improve roll and yaw control of a UAV along with drag reduction. This review also highlights the aerodynamics of flapping; specifically of the hummingbird. From the hummingbird’s aerodynamics to its hovering techniques; all of its main features are being highlighted. These birds have demonstrated a wide range of aerodynamic traits which if mimicked to near perfection could pave the way for new-age drones. Maneuverability and enhanced aerodynamic optimality could be the outset of extreme sustainability measures with these birds paving the way with their evolutionary flight measures. Apart from their aerodynamic traits, a few aspects of their wing morphing techniques and their evolutionary hereditary traits are also being highlighted.
Introduction
I. INTRODUCTION
Bird flight comprises soaring or gliding which are considered unpowered modes of flight, and flapping or hovering which are considered powered modes of flight. During soaring, birds either extract energy from the shear wind field for dynamic soaring or from thermals for thermal soaring. Hence it depends on the incident airspeed or the birds’ speed and also their genetic encoding [1]. The traits of an eagle wherein it soars for free, its wing morphing techniques, its slotted wingtips, and its efficient control surfaces that can perform roll and yaw operations with simple actuations; can help create energy-efficient Unmanned Aerial Vehicles (UAVs) with optimum aerodynamic traits. There have been few works regarding slotted wings but the incorporation of them in modern-day aircraft and their actuation remains a major challenge. This study highlights the traits of the eagle that are currently being studied by the authors to design an energy-efficient UAV with slotted wingtips requiring low actuation power. Furthermore, the flapping flight of the Hummingbird is also discussed in this work. Multiple factors of a particular niche act as agents of pressure on multiple species to evolve per se for survival. The evolution of hummingbirds is deeply rooted in the fact that their main source of energy has had its rich share of evolution, i.e. the floral nectar. The hummingbird, despite being a bird; had to evolve both anatomically and concerning locomotion to attain a flight strategy similar to that of an insect. The hummingbird while extracting nectar prefers not to stand or land but to hover. This requires a higher dissipation of energy as compared to just flying. This is also a primary reason why most of the larger birds tend to hover only with the aid of wind, thermals, etc. Hence, this strategy of the hummingbird is mostly suitable for birds of its stature and size. But the hovering strategy of a hummingbird comes with its own set of multiple challenges-For birds with a comparatively smaller stature, the up-stroke plays little role in acting as weight support hence the entire pressure of weight support falls on the bird’s down-stroke.
- This further leads to the fact that the magnitude of down-stroke must be high enough to approximately twice that of the weight effectively acting as weight support. This would further lead to vertical positional instability, i.e. within each wing-beat one would observe alternating ascents and free falls.
- Cessation of useful aerodynamic forces while executing the up-stroke maneuver would further lead to discontinuous loss of positional control concerning the other 2 dimensions.
They tend to spend a lot of their time in comparatively less dense air at higher levels of elevation.
II. LITERATURE REVIEW
The Eagle- Eagles switch between different modes of flight in response to atmospheric conditions whilst utilizing orographic and thermal updrafts. This is how these migratory raptors subsidize their flights. The distribution of thermal updrafts and orographic updrafts keeps varying as they are influenced by atmospheric perturbations and the topography of the terrain. Eagles, especially the golden eagle tend to migrate via routes where the probability to utilize thermal updrafts is higher [2].
Atmospheric conditions act as crucial factors which immensely influence any bird’s modes of flight, be it flapping or soaring. Two such influential atmospheric factors are shear-induced turbulence and updrafts. These act as outboard and external sources of energy during different soaring flight mode types. The topography of any particular region and the hindrances that it causes can also be included whilst discussing turbulent atmospheric conditions [3] [4]. Weather conditions of different terrains and regions aid in the lift generating techniques used by different bird species [5]. Some of them are-
- Orographic lift generation due to the hindrances posed by any terrain’s topographical distribution
- Lift due to energy extraction from thermal shells
Lift due to energy extraction from the shear wind field
A. Thermal Soaring and Manoeuvring
There have been multiple theories as to how birds soar. A widely accepted one is that thermal soaring is mainly due to the existence of hot air columns. In simpler terms, these columns rise from the earth’s surface at a speed higher than the speed at which the bird is sinking. Hence the bird can utilize this to attain heights [6].
The soaring of heavier vultures can be divided into two parts- first, the circling phase and, second, the gliding phase. The vulture performs, slowly yet steadily, performs multiple loops with the objective of ascending or gaining height. The vulture stretches its wing out with a bank angle inwards of the circle. Once the vulture can attain the required height, it dives and undergoes a gliding mode in a straight or linear path. It exhibits increased speed and decreased height. This cycle is carried out multiple times with the least net energy loss. Based on the patterns of the above-mentioned phases, there are also a few other types of thermal soaring. Thermal soaring is primarily influenced by the presence of thermal eddies and the intensity of atmospheric radiation, hence primarily noticed in regions with abundant solar intensity. The weight of the bird determines how much height it can attain and for how long will it loop before gliding. In a more technical sense, the wing loading of the concerned bird is the primary influence. Proper altitude is needed for efficient soaring [7].
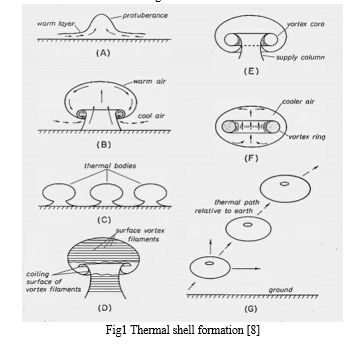
To gain more insights into the basic technical aspects of thermal soaring, Clarence D. Cone Jr. postulated that thermal soaring occurs due to a rising closed air shell and not particularly due to the continuously rising warm air columns. Now, this shell consists of a vortex core and the air in the interior region is circulating about this buoyant core. This circulatory flow is the reason behind the rising air currents. Now another reason to assume an air shell and not a continuous column is that soar-ability is efficient only in calm air, which means these “columns” do not exhibit any contact with the surface of the earth. The up-current region is the one where the bird performs the continuous loops for altitude gain. The magnitude at which this region is ascending is stronger than the magnitude of velocity at which the bird is losing altitude or sinking. Hence this enables the birds to ascend. Wind speed and direction also play a crucial role along with other factors like moisture, temperature, etc. The reason behind the assumption that circulation is exhibited inside the closed shell as opposed to a stagnant interior region is because, in a stagnant interior region, the bird will be able to ascend but would not be able to sustain the looping or circling phase. However, there exists a complex relationship between the thermal and the bird in terms of strength and aerodynamics respectively. To maintain equilibrium and sustain the looping phase, the bird has to perform circling under lower value of turn radius which subsequently increases the velocity of descent. In this case, the sinking rate will have a higher magnitude than the up-current of the thermal. These are the complexities that exist between the thermal strength of the closed shell and the aerodynamics of the bird. For this very reason, a quantitative approach is viable for a more reasonable and controlled relationship and helps explain these complexities.
The atmospheric and solar radiations heat the earth’s surface and the influence of heating is felt in the layers of air just above the surface of the earth. This is also the reason why despite the continuous solar radiation the earth’s surface does not exceed over 100 degrees. As the air above the surface along with the moisture heats up, it becomes dense as compared to the air in the layers above it, which are comparatively of lower temperature. This causes it to rise (in the shape of a protruding mushroom initially). As this moist warm air rises, its place is taken by the less dense, dry and, cool air. This thermal shell then rises due to buoyancy and is devoid of its contact with the ground surface (hence not a column of warm rising air). Like a helium balloon but without any exterior surface and with a circulation inside the shell.
This is due to the existence of vorticity which forms due to the presence of air of lower temperature outside the shell than the air temperature inside the shell [8]. Both temperature and moisture (difference in the densities of water and air) have crucial roles to play in the formation of this closed thermal shell that aids in thermal soaring. While performing a soaring maneuver, it is observable that an eagle maintains its wings at a dihedral angle (longitudinal dihedral). This in turn provides pitch stability. This also counters the nose-down pitching moment which occurs when the lift force acts aft of the raptor’s center of mass. Hence a nose-up pitching couple is observable.
To maintain the pitch stability certain wing morphing techniques are used by the eagle- sweep angle in the forward direction along with wash in wherein the wing-tips are characterized by higher values for the angle of incidence than the wing-root; sweep angle in the backward direction along with wash out wherein the wing-root is characterized by higher values for the angle of incidence than the wing-tips. Incorporating a sweep angle and an anhedral angle along with the eagle’s slotted wings is another morphing technique used by this raptor to maintain stability (yaw stability and roll stability in this case). For maintaining a stable flight within the thermal shell to perform thermal soaring maneuvers, bank turns are very crucial for the eagle. The eagle manipulates stall characteristics to make rapid changes in its yaw and roll stability. And during these banked turns, one would observe two primary characteristic tail movements. It spreads and lowers the tail during the initial bank phase with higher values of the incident and spread angles; strong tail banking in the same bank direction of the raptor wherein a positive load is acting on the tail and this helps in yaw movements for stability. Whenever the bird might feel that there is a reduction in the lift (due to low angle of attack), it uses the wing tucking maneuver. It readjusts the aerodynamic moment and lift. Even though it is still not clear if gust alleviation is the result that the bird aims for while using the wing tucking maneuver, it is the plausible answer [9]. Covert feathers are the feathers that form a covering over the other feathers for smoother airflow over the wings. During the flight, birds manipulate flow attachment and flow separation with the help of these [10].
A pictorial depiction of the formation of the thermal shell is given-
B. Control Surfaces and Aerofoil Characteristics
A leading edge high lift device that is hereditary in birds is called the Alula. This basically can be compared to a slat in an aircraft. The purpose is to increase the lift values and the values of stall angle of attack. The position and size are different for different species. Wingtips of birds are covered with strong feathers to resist aerodynamic forces and these also help in reducing the lift induced drag. Its function is similar to that of winglets in an aircraft [11]. Wingtips play a significant role in reducing the lift induced drag along with other significant parameters such as [12]-
- The take-off distance required is observed to be reduced
- Lower values of lift induced drag certainly mean that the fuel consumed by any aircraft is reduced
- And due to efficient use of fuel, the range is also seen to be increased
- High efficiency at higher altitudes
The Reynolds Number range in which birds fly is nearly similar to that of Micro Air Vehicles. The range is approximately in the range 103-105. The deflection of feathers located in the LE of the lower surface of the raptor’s arm region; resembles flaps of a conventional aircraft, especially the Krueger flap as shown in Fig.2. These feathers generally exhibit deflections when the bird undergoes unsteady maneuvers while taking off and landing; also when they encounter gusts mid-flight. The deflections occur passively as a response to the airflow acting on the feathers. The deflections exhibited by the covert feathers in the upper region are generally random whilst the lower region does exhibit some order. The degree of deflections of the covert feathers is generally higher for the angle of attacks having higher magnitudes. Striking similarities to that of a Krueger flap are observable [13]. The covert feathers of an eagle, prior to landing, act as leading edge flaps. The function is similar to the flaps used in high-performance aircraft. These feathers are deployed passively, i.e. the eagle does not utilize its muscles to flip out these feathers but instead, this is achieved due to the air or due to being lifted by the incoming flow. The Alula also works in a similar manner. The sensors when triggered act as agents to deploy these feathers when a sense of wing stall is observed [10].

The LE deflections exhibit higher fluctuations of lift coefficient than the TE deflections. The forces on the LE control surface give rise to multiple complexities of nonlinear nature. The LE deflections were crucial mostly during higher values for the angle of attack [15][16]. Experiments carried out to determine the efficiency and aerodynamic effects of these leading edge flaps have shown that the geometrical structure and the positioning of these flaps play a pivotal role during operations at high angles of attack. They act as efficient high lift generating devices at lower values of Re and they yield higher values for maximum lift. These results encourage the mimicking of these feathers and incorporating them in Micro Aerial Vehicles operating at lower values of Re. These covert feathers, whose deflections act as LE flaps tend to increase the values of Cl of the airfoil of the baseline. Rather than a high lift device, these feathers aid in the transition, i.e. they prevent the adverse effects of flight in the range of low Reynolds numbers on the laminar separation bubbles [14]. Its primary function is facilitating the bird flight at lower values of velocity at higher angles of attack; for instance, it’s highly efficient when the bird needs to land. Since we see a variance in the camber values when slats are deployed, one can observe that at higher values for angle of attack, the lift force values are increased and hence there is a delay in stalling. However, there have been multiple challenges while proposing a generalized utility of the Alula [17]. This leading edge high lift device is one of the primary reasons behind a bird’s efficient gliding mode of flight. Although there are still some clarifications to be made as to whether the deployment of the Alula is actively controlled by the bird or is dependent on the characteristics of the wind gradient. When the Alula is deployed it tends to provide a velocity field negative to that produced by the aerofoil. What this does is that it reduces the incoming wind velocity on the upper portion of the leading edge aerofoil. Hence even at higher angles of attack, due to a decrease in the higher values of the adverse pressure gradient at the upper portion of the leading edge aerofoil, stall resistant characteristics are improved. During a landing maneuver, especially observable during windy days, the bird needed to decrease its flight velocity. It does this by extending its limbs and placing them perpendicular to the path of its flight. This in turn generates more drag and lower values of velocity are observable. A decrease in velocity means the lift would be adversely affected. Hence the bird has to fly at a higher attack angle to maintain lift characteristics. Flying at higher attack angles might cause the wing to stall. In this case, the high pressure gradient (at the upper leading edge aerofoil) forces the Alula to be deployed which in turn delays the stalling produced in its vicinity.
The colour of any bird’s feathers is an indication of its strength. The colour of Alula feathers is generally darker and hence they are strong feathers. The position of the Alula is such that an adverse pressure gradient at the suction (near the upper portion of the leading edge aerofoil) would force the Alula to be deployed. The primary reason behind higher values of lift to drag ratios is that due to these wingtip feathers, whose function is similar to that of winglets in modern aircraft, lower values of lift induced drag is observed. This would in turn also cause a reduction in the values of the overall drag. The amount to which the drag values could be reduced depends on the angle of these tips as well as the number of tips involved. The positioning of these tip-sails will also determine the longitudinal static stability. The length of the tip sails is also instrumental as it affects the form drag values. At higher angles of attack, they are capable of shedding the tip vortices at faster rates. Hence, induced drag reduction rates are higher at higher angles of attack. The shedding of vortices plays a dominant role in the reduction of lift induced drag values along with the ground effect. This can be seen in soaring seabirds like the pelican and the albatross [18] [19].
The stability and control of an aircraft are adversely affected at lower values of speed. These birds generally fly at lower speeds as compared to modern-day aircraft, hence there exists an important relation between these multi-slot wings and low-speed flights other than just delaying stall. One advantage is that to vary the lateral altitude the bird does not have to make large movements such as is seen in fixed-wing aircrafts. For example, if the bird needs to achieve a bank angle, unlike an aircraft that would have to alter the lift on either portside or starboard to do this, the bird would instead make small movements with less dissipation of energy to achieve this. This is due to the existence of feathers at the wingtips and the leading edge slots [20] [16]. This also helps the birds in an efficient flight at lower speeds as at lower values of speed; these birds generally fly at higher angle of incidence. The existence of slots at the tips of the wing also ensures that birds do not require twisting their entire wings. Birds can also manipulate the wing sweep to increase the critical Mach number and for span-wise lift flow. This eliminates the formation of shock waves during subsonic diving maneuver; hence they can fly at lower speeds with the same amount of lift. The airfoils of eagles are different from that of modern day aircrafts. The high camber exhibited by the raptor does resemble the airfoils widely used in the aerospace industry for the purpose of obtaining higher lift values during operations at low Reynolds numbers. The raptor’s wing exhibits a wash-in distribution which implies that it exhibits a forward sweep and a positive twist from wing-root to wing-tip. The LE is thick while the TE is thinner and exhibits high degrees of flexibility which facilitates passive/automated camber manipulations. This combined with high camber properties has proven to be quite crucial when the raptor’s wing is under positive aerodynamic loading. This facilitates complex maneuvering wherein during instances of pitching up; the wing undergoes pitching and morphing. Flow separation causes the deflections on the upper region of covert feathers. The raptor’s airfoil exhibits a wide range for the values of the angle of attack and exhibits low values of aerodynamic drag. The escape speed of birds is also highly affected by their wing loading. It is inversely proportional to wing loading. Effects of gender and year etc are comparatively less prominent [21][22].
The Hummingbird-

Fig. 3 Hummingbird
For any natural flyer, the muscles primarily responsible for the down-stroke are the pectoralis and the ones involved in the up-stroke are the supracoracoideus; which in the case of hummingbirds is almost half in size as compared to the pectoralis. Almost half the volume of the supracoracoideus contains mitochondria to sustain the high frequency of wing beats and higher values of aerobic power. The hummingbird’s wing is characterized by higher values of aspect ratio and is smaller in size, narrow, and exhibits a thin leading edge which is instrumental in providing higher lift values as compared to round leading edges.

Fig.4 Hummingbird wing model [23]
On the contrary, A Non-flapping bird uses potential energy and then converts it to counter aerodynamic forces. The bird moves in two directions i.e. it moves forward in the horizontal direction and it also falls/sinks down.
Sinking rate of bird is greatly affected by three factors-
- Weight of the wing
- Morphology of the wing
- Shape of the body
Each bird uses flapping and gliding patterns in its routine, but birds can only incorporate these both patterns with the help of morphing.
a. Gliding
- High aspect ratio and elliptical platform
- Flow of wing is uniform on leading edge
b. Flapping
- Movement of tip of the wing is faster than root of the wing
- Velocity gradient is produced in the direction of wingspan
But this can be balanced with the help of tapered wing or twisted wing.
A. Aerodynamics
Due to their adaptations concerning their musculoskeletal form, hummingbirds can produce 25-33% of their body weight during up-stroke. But the most remarkable quality is the hummingbird’s ability to manipulate the fluid flow with optimum precision during the upstroke-downstroke transitions. As mentioned the sharp leading edge does provide better aerodynamic performance than a rounded one. The circulation around the wing is primarily due to the difference in pressure created due to the sharp leading edge which further results in the generation of a vortex. This vortex then undergoes circulation around the chord. The hummingbird could thus rotate its wing (around the long axis) within the vortex itself without actually interrupting or disturbing it. The translational movements exhibited by the hummingbird’s wing produces aerodynamic forces which are sustained by the bird as the wings are pulled back to their initial position to re-initiate the cycle again. In case of any interruptions which might occur whilst produce these aerodynamic forces due to multiple reasons, these can be compensated by stabilizing the bird’s head position which it effectively does via the motion of its neck [24] [25].
Despite operating at different values of Re; it was speculated that there existed similar kinematic approaches between hummingbirds and insects. With the help of flow visualization and other methods, this speculation was proven to be somewhat correct. The lift generation in hummingbirds had striking similarities to those exhibited by insects. The wing undergoing translation had leading edge vorticity (LEV) attached to its dorsal region and this was primarily the reason behind the generation of lift. Also as mentioned earlier, most of the lift used for weight support was generated due to the down-stroke rather than the up-stroke and the value comes out to be around 75%. The positive camber exhibited by the bird was primarily the reason behind this asymmetrical distribution in lift generation and also its inability to reverse during the up-stroke. The airfoil of a hummingbird also acts as a dynamic stalling airfoil [26] [1].
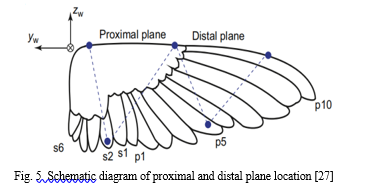
For the purpose of extraction of the involved wing kinematics, anatomical landmarks are located in the form of proximal plane and the distal planes on the primary feathers as well as the secondary ones.
The elliptical shape is suitable for subsonic flow as it creates less amount of drag and heat generation. Von Karman is suitable for supersonic flow as it creates weak shockwaves.
The wing shape of the hummingbird is not static and hence the varying dynamic wing shape and size add to the complexity of the aerodynamic forces. The Principle components patterns can be confirmed through the examination of the wing area time course. During the stroke, the wing area is increased until reached a maximum mid downstroke, and as the approach of the wing is supinated it decreases. Due to the activity of secondary feathers, there is variation in wing area while in the stroke cycle. Compression in secondary feathers causes the wing area to be reduced in upstroke independent of the wing length. The secondary feathers are continuously pressed against the body during upstroke followed by the secondaries fanning out before pronation. The dynamic behavior of the wing is characterized in accordance with the aerodynamic variables twist and camber.
Linear twisting over the wingspan in the process of mid-downstroke and mid-upstroke is exhibited by the wings according to the aerodynamic theory.
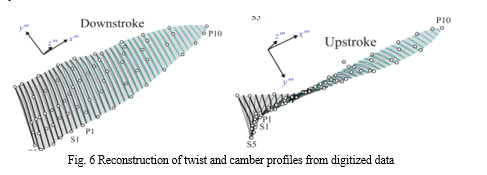
During the downstroke wing camber is 10-15% smaller. The handwing twisting is similar to deformed wing surface and hence it is difficult to reconstruct the ventral wing surface. Spanwise cambering is occurred during both the strokes ; In downstroke it is produced due to tip drag, while in upstroke it is produced because the wing is being cupped in the direction opposite to its propagation [28].
B. Wake
The wake of the hummingbird is more complex in nature while hovering. It is possible to have distinct wakes of different species. The study suggests that 64% of the supported weight is produced while downstroke with a tiny amount of differences in the kinematics of the phases in strokes. On an average, 66% of weight support is seen in the Anna’s hummingbird. The wings of Anna’s hummingbird are characterized by the formation of individual ring-like structures during each individual stroke. This kind of wake is further characterized by separate vortex loops for both the hummingbird’s wings which can be seen to shed towards the end of downstroke and travel in the ventral direction. On the other hand, during upstroke the shed loops travel dorsally [29].
Separate stop vortices are seen during the downstroke and started vortices are seen during the bird’s upstroke maneuver. Apart from this, a separate start/stop vortex can be observed at the beginning of the upstroke and end of downstroke. These transitions are quick between the stroke phases which combines separate vortex structures and hence it results in complex wake [30] [19].
Once the down-stroke is initiated for translation, one would observe circulation resulting from the up-stroke shed. The far-field wake is characterized by the production of only 1 shed vortex at the upstroke-downstroke transition. This transition is so smooth or that the up-stroke stopping vortex is extremely close spatially with the down-stroke starting vortex that they almost seem to merge. But the situation is different in the case of the vortex shed at the down-stroke ending point where it is convected down due to the greater magnitude of downwash produced by the down-stroke. This leads to the generation of a down-stroke stopping vortex distinct from the up-stroke starting vortex.
C. Bound and Leading Edge Vorticity Circulation
The bound and leading-edge vorticity (LEV) circulation exhibited by a hummingbird’s wing is comparatively different from that exhibited by different insects, where one would observe flow separation of higher magnitude and retrograde. The lift generation is highly affected by the low-pressure region at the sharp leading edge as a bound circulation is created which is quite sufficient to generate the adequate lift required. The fluid flow from the ventral region gets drawn and used for the creation of this bound circulation with laminar properties which further encompasses the chord [31].

Fig. 7 (a) Schematic depicting the structure of simplest valid LEV for a cylindrical vortex—cross-section view (b) An idealized top view schematic depicting LEV of conical shape and its topology for wing that undergoes steady revolution with a focus at its root.[32]
Leading Edge Vortex is the firm solution for preventing stalling. The wake study of the hummingbird shows that during downstroke at lower Reynold’s number LEV dominates. LEV is used as the lift mechanism because they instead of shedding into the wake as the lift is lost while stalling are stable and attach to the wing during translation. The LEV is possibly created by stable flows at low Reynolds number [29].
As studied by Warrick, when hummingbird is characterized by aerodynamics exhibited at the wing’s sharp leading edge, LEV may vary between 0.7% to 26% of total mass production. In addition to this, the LEV is always created within 3mm of the wing’s dorsal surface [33].
D. Rotation of Wing
As discussed earlier this bound circulation is primarily the reason behind the production of aerodynamic forces. The hummingbird’s wing has a unique ability to sustain this produced bound circulation without the shed of vortices.
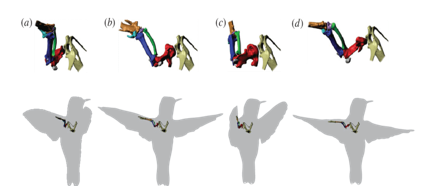
Fig. 8 Estimation of the orientation and position of the bird’s shoulder and proximal forelimb bones while performing a a complete stroke cycle during hovering: (a) during the initiation of downstroke, (b) mid-downstroke, (c) end of downstroke, (d) middle of upstroke [34]
E. Hovering
Hovering manoeuvre is carried out by the bird via generation of stable LEV that further enables the hummingbird to sustain stability at higher angles of attack with aid of delayed stall. Flow in the root’s proximity is observed to be attached fully and separation of the shear layer is observed to occur at around 75% of the hummingbird’s wing-span where it exhibits a connection with the tip vortex. Even though flow field studies show a significant symmetry for both the up-stroke and down-stroke, it is quite necessary to take into account aerodynamic parameters as well. As already discussed the lift contribution of the down-stroke is comparatively higher than the up-stroke. Here a reduced value of drag is also observed as compared to the up-stroke drag generation although there are multiple variables to be taken into account during drag determination such as the role of the musculoskeletal actuation. But in general terms, the drag is comparatively lower during the down-stroke. Unlike insects, the hummingbird’s wing exhibits a camber profile. This facilitates a suction force in the forward direction near the LE induced by the LEV. The evolution of hummingbirds and their genetic encodings have enabled them to exhibit enhanced hover performances by flapping their wings during the stroke; at AoA which are sub-optimal in terms of aerodynamics. This not only helps the bird to minimize the inertial losses but also enables the bird to have optimum utilization of muscle mechanics during the bird’s forward and hovering flight. Another point of importance is that hummingbirds exhibit higher values of aspect ratio and tend to require comparatively less power to sustain and support their body weight than birds who exhibit wings with comparatively lower values of aspect ratio. The hummingbird’s wings are optimal for hovering and if mimicked for modern-day helicopters than optimal slow hovering flight performance manoeuvrability can be facilitated [35].
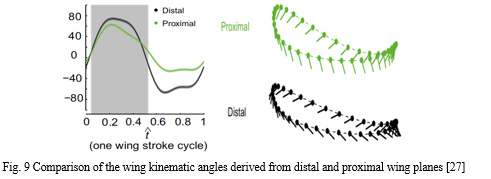
The hummingbirds are evolved over a period of time with an aerodynamic mechanism and are capable of hovering by high frequency and with wing motions at higher values of angle of attack. The kinematics derived from the distal wing planes and the proximal planes have a great amount of difference concerning the rotational angles particularly in the case of the upstroke. When flapping, wings undergo large spanwise twists and minor spanwise bending.
Previous studies on hovering hummingbirds show that wing deformation includes chordwise camber and spanwise twist. The upstroke is characterized by a more substantial spanwise twist as compared to the downstroke.
The pattern of wing twists produced by a hummingbird during hovering exhibits similarities to the “washout” design incorporated in helicopter blades and propellers. This design helps in reducing the induced power loss and thus increases the aerodynamic efficiency [36].
III. ACKNOWLEDGMENT
The authors thank the Department of Aerospace Engineering, Lovely Professional University, Phagwara for their guidance and support in the successful completion of this work.
Conclusion
The Emergence of bio-mimetic drones has paved the way for more efficient and aerodynamic drones in multiple sectors. Analyzing the aerodynamic efficiencies and the structural composition of the eagle’s wing along with its soar-ability can be instrumental in creating energy-efficient drones with minimal energy dissipation and with more complex yet far more superior control surfaces. These findings are studied by the authors for the design and construction of bionic wingtips and a wing with bell shaped lift distribution providing superior roll control with less actuation power. Another aim is to eliminate the sweep in tail-less UAVs without affecting stability. The review also shed light on the different traits of the hummingbird’s flapping aerodynamics and drew parallels to modern aerodynamics. The hovering techniques used by the hummingbirds helps in achieving enhanced aerodynamic efficiency but at a cost in terms of energy. Similar parallels can be drawn for a helicopter as well. As discussed, this washout technique when incorporated in the propeller blades enhances the aerodynamic efficiency. Similarly flapping and rotational techniques used by hummingbirds can aid in achieving better slow hovering flight performance in modern day helicopters. The delayed stall technique discussed earlier also aims at achieving improved stability at higher angles of attack. The complex and distinct wake formations and the wake study can confirm this statement. Mimicking the hummingbird’s traits can help improve performance of modern day aircrafts.
References
[1] B. C. Mathew, J. V. M. Lal Jeyan, P. Dutta, and R. R. Savale, “Evolutionary and Hereditary Traits of an Albatross and its Aerodynamic Optimality,” IOP Conf. Ser. Earth Environ. Sci., vol. 775, no. 1, p. 012002, Jun. 2021, doi: 10.1088/1755-1315/775/1/012002. [2] T. E. Katzner et al., “Use of multiple modes of flight subsidy by a soaring terrestrial bird, the golden eagle Aquila chrysaetos, when on migration,” J. R. Soc. Interface, vol. 12, no. 112, Nov. 2015, doi: 10.1098/rsif.2015.0530. [3] J. M. Mallon, K. L. Bildstein, and T. E. Katzner, “In-flight turbulence benefits soaring birds,” Auk, vol. 133, no. 1, pp. 79–85, Jan. 2016, doi: 10.1642/AUK-15-114.1. [4] K. M. Laurent et al., “Turbulence explains the accelerations of an eagle in natural flight,” Proc. Natl. Acad. Sci., vol. 118, no. 23, Jun. 2021, doi: 10.1073/PNAS.2102588118. [5] A. E. Duerr et al., “Flight response of slope-soaring birds to seasonal variation in thermal generation,” Funct. Ecol., vol. 29, no. 6, pp. 779–790, Jun. 2015, doi: 10.1111/1365-2435.12381. [6] Z. Ákosl, M. Nagy, S. Leven, and T. Vicsek, “Thermal soaring flight of birds and unmanned aerial vehicles,” Bioinspiration and Biomimetics, vol. 5, no. 4, Dec. 2010, doi: 10.1088/1748-3182/5/4/045003. [7] H. J. Williams, O. Duriez, M. D. Holton, G. Dell’Omo, R. P. Wilson, and E. L. C. Shepard, “Vultures respond to challenges of near-ground thermal soaring by varying bank angle,” J. Exp. Biol., vol. 221, no. 23, Dec. 2018, doi: 10.1242/JEB.174995. [8] “THERMAL SOARING OF BIRDS on JSTOR.” https://www.jstor.org/stable/27838346?seq=1 (accessed May 04, 2021). [9] K. V. Reynolds, A. L. R. Thomas, and G. K. Taylor, “Wing tucks are a response to atmospheric turbulence in the soaring flight of the steppe eagle Aquila nipalensis,” J. R. Soc. Interface, vol. 11, no. 101, Dec. 2014, doi: 10.1098/rsif.2014.0645. [10] A. C. Carruthers, A. L. R. Thomas, and G. K. Taylor, “Automatic aeroelastic devices in the wings of a steppe eagle Aquila nipalensis,” J. Exp. Biol., vol. 210, no. 23, pp. 4136–4149, Dec. 2007, doi: 10.1242/jeb.011197. [11] Y. Murayama, T. Nakata, and H. Liu, “Flexible Flaps Inspired by Avian Feathers Can Enhance Aerodynamic Robustness in low Reynolds Number Airfoils,” Front. Bioeng. Biotechnol., vol. 0, p. 374, May 2021, doi: 10.3389/FBIOE.2021.612182. [12] N. A. Siddiqui, “Experimental investigation on the effect of biomimetic wingtips on aerodynamics of wings,” 2018, Accessed: Sep. 11, 2021. [Online]. Available: http://studentrepo.iium.edu.my/handle/123456789/4799. [13] A. C. Carruthers, G. K. Taylor, S. M. Walker, and A. L. R. Thomas, “Use and function of a leading edge flap on the wings of eagles,” in Collection of Technical Papers - 45th AIAA Aerospace Sciences Meeting, 2007, vol. 1, pp. 382–390, doi: 10.2514/6.2007-43. [14] N. M. Bakhtian, H. Babinsky, A. L. R. Thomas, and G. K. Taylor, “The low Reynolds number aerodynamics of leading edge flaps,” in Collection of Technical Papers - 45th AIAA Aerospace Sciences Meeting, 2007, vol. 12, pp. 8018–8030, doi: 10.2514/6.2007-662. [15] A. Panta et al., “Low Reynolds number aerodynamics of leading-edge and trailing-edge hinged control surfaces: Part I statics,” Aerosp. Sci. Technol., vol. 99, p. 105563, Apr. 2020, doi: 10.1016/J.AST.2019.105563. [16] P. Dutta and A. Roshan, “Bell Shaped Lift Distribution, Aerodynamic Soaring and Control Surface Dynamics of Birds,” Int. J. Res. Appl. Sci. Eng. Technol., vol. 10, no. 8, pp. 184–189, Aug. 2022, doi: 10.22214/IJRASET.2022.46134. [17] S. I. Lee, J. Kim, H. Park, P. G. Jab?o?ski, and H. Choi, “The function of the alula in avian flight,” Sci. Rep., vol. 5, no. 1, pp. 1–5, May 2015, doi: 10.1038/srep09914. [18] J. xun Zhou, C. hong Sun, and Daichin, “Drag reduction and flow structures of wing tip sails in ground effect,” J. Hydrodyn., vol. 32, no. 1, pp. 93–106, Feb. 2020, doi: 10.1007/s42241-020-0006-4. [19] B. C. Mathew, S. K. Sahu, P. Dutta, R. Savale, and M. Jv, “Issue 3 Article 1 Part of the Aerodynamics and Fluid Mechanics Commons, and the Aeronautical Vehicles Commons Scholarly Commons Citation Scholarly Commons Citation,” Int. J. Aviat. Aeronaut. Aerosp., vol. 8, no. 3, 2021, doi: 10.15394/ijaaa.2021.1598. [20] P. Dutta, O. P. Nagar, S. K. Sahu, R. R. Savale, and R. Gokul Raj, “Aerodynamic analysis of bionic winglet- slotted wings,” Mater. Today Proc., vol. 62, pp. 6701–6707, Jan. 2022, doi: 10.1016/J.MATPR.2022.04.752. [21] J. W. Lish, R. Domenech, B. E. Bedrosian, D. H. Ellis, and M. Payton, “Wing Loading in North American Golden Eagles Aquila chrysaetos,” J. Raptor Res., vol. 50, no. 1, pp. 70–75, Mar. 2016, doi: 10.3356/rapt-50-01-70-75.1. [22] J. G. Burns and R. C. Ydenberg, “The effects of wing loading and gender on the escape flights of least sandpipers (Calidris minutilla) and western sandpipers (Calidris mauri),” Behav. Ecol. Sociobiol., vol. 52, no. 2, pp. 128–136, Jul. 2002, doi: 10.1007/s00265-002-0494-y. [23] M. Maeda, T. Nakata, I. Kitamura, H. Tanaka, and H. Liu, “Quantifying the dynamic wing morphing of hovering hummingbird,” R. Soc. Open Sci., vol. 4, no. 9, Sep. 2017, doi: 10.1098/rsos.170307. [24] D. Warrick, T. Hedrick, M. J. Fernández, B. Tobalske, and A. Biewener, “Hummingbird flight,” Current Biology, vol. 22, no. 12. Cell Press, pp. R472–R477, Jun. 19, 2012, doi: 10.1016/j.cub.2012.04.057. [25] D. R. Warrick, B. W. Tobalske, and D. R. Powers, “Aerodynamics of the hovering hummingbird,” Nature, vol. 435, no. 7045, pp. 1094–1097, Jun. 2005, doi: 10.1038/nature03647. [26] D. R. Warrick, B. W. Tobalske, D. R. Powers, and M. H. Dickinson, “The aerodynamics of hummingbird flight,” in Collection of Technical Papers - 45th AIAA Aerospace Sciences Meeting, 2007, vol. 1, pp. 367–371, doi: 10.2514/6.2007-41. [27] B. Cheng et al., “Flight mechanics and control of escape manoeuvres in hummingbirds. I. Flight kinematics,” J. Exp. Biol., vol. 219, no. 22, pp. 3518–3531, Nov. 2016, doi: 10.1242/JEB.137539. [28] D. A. Skandalis, “Form and function in hummingbird flight,” 2017, doi: 10.14288/1.0361753. [29] Y. Nan, L. Chen, D. Mcglinchey, B. Peng, Y. I. Chen, and Y. Chen, “Smart Robot Carer View project Computational Intelligence Aided Design and Manufacture View project SPECIAL SECTION ON UNCERTAINTY QUANTIFICATION IN ROBOTIC APPLICATIONS From Studying Real Hummingbirds to Designing Hummingbird-Like Robots-A Literature Review,” doi: 10.1109/ACCESS.2019.2939491. [30] D. R. Warrick, B. W. Tobalske, and D. R. Powers, “Aerodynamics of the hovering hummingbird,” Nat. 2005 4357045, vol. 435, no. 7045, pp. 1094–1097, Jun. 2005, doi: 10.1038/nature03647. [31] D. R. Warrick, B. W. Tobalske, and D. R. Powers, “Lift production in the hovering hummingbird,” Proc. R. Soc. B Biol. Sci., vol. 276, no. 1674, pp. 3747–3752, Nov. 2009, doi: 10.1098/rspb.2009.1003. [32] M. R. A. Nabawy and W. J. Crowther, “The role of the leading edge vortex in lift augmentation of steadily revolving wings: A change in perspective,” Journal of the Royal Society Interface, vol. 14, no. 132. Royal Society Publishing, Jul. 01, 2017, doi: 10.1098/rsif.2017.0159. [33] D. R. Warrick, B. W. Tobalske, and D. R. Powers, “Lift production in the hovering hummingbird,” Proc. R. Soc. B Biol. Sci., vol. 276, no. 1674, pp. 3747–3752, Nov. 2009, doi: 10.1098/RSPB.2009.1003. [34] T. L. Hedrick, B. W. Tobalske, I. G. Ros, D. R. Warrick, and A. A. Biewener, “Morphological and kinematic basis of the hummingbird flight stroke: Scaling of flight muscle transmission ratio,” Proc. R. Soc. B Biol. Sci., vol. 279, no. 1735, pp. 1986–1992, May 2012, doi: 10.1098/rspb.2011.2238. [35] J. W. Kruyt, E. M. Quicazán-Rubio, G. F. Van Heijst, D. L. Altshuler, and D. Lentink, “Hummingbird wing efficacy depends on aspect ratio and compares with helicopter rotors,” J. R. Soc. Interface, vol. 11, no. 99, Oct. 2014, doi: 10.1098/rsif.2014.0585. [36] “Principles of Helicopter Aerodynamics with CD Extra - Gordon J. Leishman - Google Books.” https://books.google.co.in/books?hl=en&lr=&id=nMV-TkaX-9cC&oi=fnd&pg=PR19&dq=Principles+of+Helicopter+Aerodynamics.+New+York:+Cambridge+University+Press.&ots=CsaSkbXIaC&sig=6afTIb0-komVd_OuXFMPS8A4thc&redir_esc=y#v=onepage&q=Principles of Helicopter Aerodynamics. New York%3A Cambridge University Press.&f=false (accessed Oct. 17, 2021).
Copyright
Copyright © 2022 Prantik Dutta, Samruddhi Chaugule. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

Download Paper
Paper Id : IJRASET47148
Publish Date : 2022-10-20
ISSN : 2321-9653
Publisher Name : IJRASET
DOI Link : Click Here
 Submit Paper Online
Submit Paper Online